Konference: 2005 XXIX. Brněnské onkologické dny a XIX. Konference pro sestry a laboranty
Kategorie: Nádorová biologie/imunologie/genetika a buněčná terapie
Téma: Pokroky v molekulární biologii nádorů
Číslo abstraktu: 043
Autoři: prof. RNDr. Jana Šmardová, CSc.; Mgr. Šárka Pavlová, PhD; Miluše Svitáková; Mgr. Diana Grochová; RNDr. Barbora Ravčuková
Úvod
Nádorový supresor p53 je sekvenčně specifický transkripční faktor, který se aktivuje v buňkách vystavených stresovým stavům a následně ovlivňuje expresi svých cílových genů. Gen p53 je často mutován v buňkách lidských nádorů a nádorových linií. K detekci mutací p53 lze použít několika různých metod, mezi které patří také funkční analýza separovaných alel v kvasinkách (FASAY). Tato metoda je založena jednak na existenci kvasinkového reparačního mechanismu na principu homologní rekombinace („gap repair“) a jednak na transkripčně aktivační schopnosti proteinu p53 v kvasinkových buňkách. Protein p53 vytvořený reverzně transkripční PCR z mRNA vyšetřované tkáně a následnou translací v kvasinkách je podroben analýze svých transkripčně aktivačních schopností s využitím reportérského genu ADE2. Míra transaktivační schopnosti p53 je hodnocena podle zbarvení kvasinkových kolonií, které rostou na plotnách s nízkou hladinou adeninu a které exprimují danou variantu p53 (1). FASAY je semikvantitativní metoda, která navíc dokáže rozlišit plně inaktivující mutace p53 od mutací pouze částečně snižujících transaktivační schopnost proteinu p53. Využitím dalších variant metody FASAY lze analyzovat i detailnější funkční rozdíly mezi jednotlivými typy mutací p53 (2). Právě tato funkční metoda v posledních letech významně přispěla k objevům velké rozmanitosti mutantů p53 a vedla k formulaci modelu „p(piano)53“, který vyjadřuje moderní pohled na variabilitu funkčních dopadů různých mutací p53 (3). Metoda FASAY se osvědčila také v klinických studiích při analýze statutu p53 v různých tkáních, protože detekuje mutace p53 s vysokou přesností a citlivostí. V této práci jsme metodou FASAY vyšetřili status p53 u 26 buněčných linií odvozených z různých typů lidských nádorů. Získané výsledky zřetelně demonstrují přednosti metody FASAY pro klinické aplikace. Jedním z našich významných objevů byla identifikace unikátní dosud nepopsané „non-sense“ mutace genu p53 v kodónu 124.
Výsledky
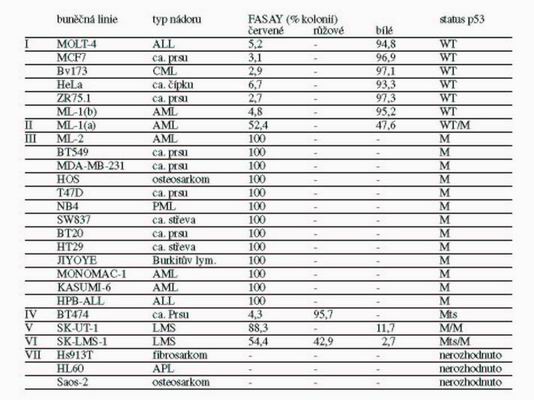
Funkčním testem v kvasinkách jsme analyzovali status p53 u 26 buněčných linií odvozených z různých typů lidských nádorů a na jeho základě jsme tyto linie rozdělili do sedmi skupin (Tabulka 1). Linie MOLT-4, MCF7, Bv173, HeLa, ZR75.1 a ML-
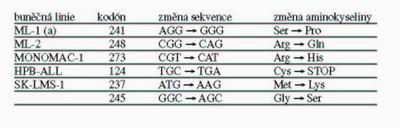
1(b), u kterých podíl červených kolonií nepřesahoval 10 %, tedy hodnotu pozadí metody, byly zařazeny do I. skupiny a jejich status p53 byl považován za standardní (wt). Buněčná linie ML-1(a) tvořila II. skupinu, které FASAY přisoudila heterozygotní stav p53, protože frekvence červených kolonií dosáhla 52,4 %. III. skupinu tvořily buněčné linie BT549, MDA-MB-231, HOS, T47D, NB4, SW837, BT20, HT29, JIYOYE, KASUMI-6, ML-2, MONOMAC-1 a HPB-ALL, které nesly pouze nefunkční alely p53: FASAY prokázala 100 % podíl červených kolonií. U linií, u kterých byl status p53 dosud neznámý, jsme nově zachycené mutace analyzovali sekvenováním DNA (Tabulka 2). Mezi liniemi s mutantními alelami představovala linie BT474 specifický případ a tvořila skupinu IV., přestože výsledkem FASAY bylo také 100 % jiných než bílých kolonií jako u linií ze III. skupiny. V případě linie BT474 bylo ale typicky červených pouze 4,3 % kolonií, které představovaly pozadí metody, ostatní kolonie byly růžové, což svědčilo o teplotně senzitivním charakteru mutace p53 v buňkách BT474. Také u linií SK-UT-1 a SK-LMS-1, které nesou mutantní p53, jsme očekávali 100 % podíl kolonií s jiným než bílým zbarvením. U linie SK-UT-1 (skupina V.) byla prokázána přítomnost dvou bodových mutací (v kodónech 175 a 248), z nich každá pravděpodobně postihuje jinou alelu genu p53. Výsledkem FASAY bylo ovšem pouze 88,3 % červených kolonií a 11,7 % bílých kolonií. Poměr červených a bílých kolonií zůstával stabilní i po opakovaných pasážích buněk v kultuře. 11,7 % červených kolonií zřejmě představuje frekvenci rekombinace, ke které dochází uvnitř buněk mezi alelami genu p53. Skupinu VI. tvoří linie SK-LMS-1. Výsledkem analýzy buněk této linie metodou FASAY bylo 54,4 % červených kolonií, 42,9 % růžových kolonií a 3 % bílých kolonií. Přítomnost inaktivující mutace v kodónu 245 byla popsána již dříve, ale náš výsledek svědčil o přítomnosti další dosud neznámé teplotně senzitivní mutaci v druhé alele genu p53. Tento předpoklad jsme potvrdili sekvenováním DNA (Tabulka 2). Nízké procento bílých kolonií opět svědčilo o přítomnosti alel bez mutace p53, které pravděpodobně vznikly rekombinací uvnitř buněk SK-LMS-1. Nižší frekvence rekombinace ve srovnání se vzorky linie SK-UT-1 odpovídala kratší vzdálenosti mezi nalezenými mutacemi (237-245 u SK-LMS-1 versus 175-248 u SK-UT-1). Poslední, VII. skupinu tvořily buněčné linie Hs913T, HL60 a Saos-2, u kterých FASAY nedokázala status p53 jednoznačně určit v důsledku přítomnosti rozsáhlých delecí a přestaveb genu p53.
U vybraných buněčných linií jsme rovněž provedli analýzu hladiny proteinu p53 imunoblotingem s využitím protilátky DO-
1. V buňkách ML-1(a), ML-1(b), ML-2 a MONOMAC-1 jsme detekovali vysokou hladinu proteinu p53. V buňkách HPB-ALL, ve kterých FASAY prokázala přítomnost mutace p53, jsme překvapivě protein p53 nedetekovali. Analýzou sekvence DNA jsme tento překvapivý výsledek vysvětlili nálezem „non-sense“ mutace v kodónu 124 genu p53, která vede k předčasnému ukončení translace (Tabulka 2). V databázi IARC (http://www.iarc.fr/p53) (4) je v současné době (1/2005) zaznamenáno 19 806 mutací, ale mutace 124STOP mezi nimi není. Jedná se pravděpodobně o zcela novou mutaci, která dosud nebyla popsána.
Závěry
Potvrdili jsme, že FASAY je velmi spolehlivá metoda pro stanovení funkčního stavu proteinu p53. U buněčných linií, jejichž status p53 byl popsán již dříve, jsme předchozí výsledek buď potvrdili nebo zpřesnili. V několika případech nám FASAY poskytla hlubší vhled do povahy statutu p53.
Nádorový supresor p53 je sekvenčně specifický transkripční faktor, který se aktivuje v buňkách vystavených stresovým stavům a následně ovlivňuje expresi svých cílových genů. Gen p53 je často mutován v buňkách lidských nádorů a nádorových linií. K detekci mutací p53 lze použít několika různých metod, mezi které patří také funkční analýza separovaných alel v kvasinkách (FASAY). Tato metoda je založena jednak na existenci kvasinkového reparačního mechanismu na principu homologní rekombinace („gap repair“) a jednak na transkripčně aktivační schopnosti proteinu p53 v kvasinkových buňkách. Protein p53 vytvořený reverzně transkripční PCR z mRNA vyšetřované tkáně a následnou translací v kvasinkách je podroben analýze svých transkripčně aktivačních schopností s využitím reportérského genu ADE2. Míra transaktivační schopnosti p53 je hodnocena podle zbarvení kvasinkových kolonií, které rostou na plotnách s nízkou hladinou adeninu a které exprimují danou variantu p53 (1). FASAY je semikvantitativní metoda, která navíc dokáže rozlišit plně inaktivující mutace p53 od mutací pouze částečně snižujících transaktivační schopnost proteinu p53. Využitím dalších variant metody FASAY lze analyzovat i detailnější funkční rozdíly mezi jednotlivými typy mutací p53 (2). Právě tato funkční metoda v posledních letech významně přispěla k objevům velké rozmanitosti mutantů p53 a vedla k formulaci modelu „p(piano)53“, který vyjadřuje moderní pohled na variabilitu funkčních dopadů různých mutací p53 (3). Metoda FASAY se osvědčila také v klinických studiích při analýze statutu p53 v různých tkáních, protože detekuje mutace p53 s vysokou přesností a citlivostí. V této práci jsme metodou FASAY vyšetřili status p53 u 26 buněčných linií odvozených z různých typů lidských nádorů. Získané výsledky zřetelně demonstrují přednosti metody FASAY pro klinické aplikace. Jedním z našich významných objevů byla identifikace unikátní dosud nepopsané „non-sense“ mutace genu p53 v kodónu 124.
Výsledky
Funkčním testem v kvasinkách jsme analyzovali status p53 u 26 buněčných linií odvozených z různých typů lidských nádorů a na jeho základě jsme tyto linie rozdělili do sedmi skupin (Tabulka 1). Linie MOLT-4, MCF7, Bv173, HeLa, ZR75.1 a ML-
1(b), u kterých podíl červených kolonií nepřesahoval 10 %, tedy hodnotu pozadí metody, byly zařazeny do I. skupiny a jejich status p53 byl považován za standardní (wt). Buněčná linie ML-1(a) tvořila II. skupinu, které FASAY přisoudila heterozygotní stav p53, protože frekvence červených kolonií dosáhla 52,4 %. III. skupinu tvořily buněčné linie BT549, MDA-MB-231, HOS, T47D, NB4, SW837, BT20, HT29, JIYOYE, KASUMI-6, ML-2, MONOMAC-1 a HPB-ALL, které nesly pouze nefunkční alely p53: FASAY prokázala 100 % podíl červených kolonií. U linií, u kterých byl status p53 dosud neznámý, jsme nově zachycené mutace analyzovali sekvenováním DNA (Tabulka 2). Mezi liniemi s mutantními alelami představovala linie BT474 specifický případ a tvořila skupinu IV., přestože výsledkem FASAY bylo také 100 % jiných než bílých kolonií jako u linií ze III. skupiny. V případě linie BT474 bylo ale typicky červených pouze 4,3 % kolonií, které představovaly pozadí metody, ostatní kolonie byly růžové, což svědčilo o teplotně senzitivním charakteru mutace p53 v buňkách BT474. Také u linií SK-UT-1 a SK-LMS-1, které nesou mutantní p53, jsme očekávali 100 % podíl kolonií s jiným než bílým zbarvením. U linie SK-UT-1 (skupina V.) byla prokázána přítomnost dvou bodových mutací (v kodónech 175 a 248), z nich každá pravděpodobně postihuje jinou alelu genu p53. Výsledkem FASAY bylo ovšem pouze 88,3 % červených kolonií a 11,7 % bílých kolonií. Poměr červených a bílých kolonií zůstával stabilní i po opakovaných pasážích buněk v kultuře. 11,7 % červených kolonií zřejmě představuje frekvenci rekombinace, ke které dochází uvnitř buněk mezi alelami genu p53. Skupinu VI. tvoří linie SK-LMS-1. Výsledkem analýzy buněk této linie metodou FASAY bylo 54,4 % červených kolonií, 42,9 % růžových kolonií a 3 % bílých kolonií. Přítomnost inaktivující mutace v kodónu 245 byla popsána již dříve, ale náš výsledek svědčil o přítomnosti další dosud neznámé teplotně senzitivní mutaci v druhé alele genu p53. Tento předpoklad jsme potvrdili sekvenováním DNA (Tabulka 2). Nízké procento bílých kolonií opět svědčilo o přítomnosti alel bez mutace p53, které pravděpodobně vznikly rekombinací uvnitř buněk SK-LMS-1. Nižší frekvence rekombinace ve srovnání se vzorky linie SK-UT-1 odpovídala kratší vzdálenosti mezi nalezenými mutacemi (237-245 u SK-LMS-1 versus 175-248 u SK-UT-1). Poslední, VII. skupinu tvořily buněčné linie Hs913T, HL60 a Saos-2, u kterých FASAY nedokázala status p53 jednoznačně určit v důsledku přítomnosti rozsáhlých delecí a přestaveb genu p53.
U vybraných buněčných linií jsme rovněž provedli analýzu hladiny proteinu p53 imunoblotingem s využitím protilátky DO-
1. V buňkách ML-1(a), ML-1(b), ML-2 a MONOMAC-1 jsme detekovali vysokou hladinu proteinu p53. V buňkách HPB-ALL, ve kterých FASAY prokázala přítomnost mutace p53, jsme překvapivě protein p53 nedetekovali. Analýzou sekvence DNA jsme tento překvapivý výsledek vysvětlili nálezem „non-sense“ mutace v kodónu 124 genu p53, která vede k předčasnému ukončení translace (Tabulka 2). V databázi IARC (http://www.iarc.fr/p53) (4) je v současné době (1/2005) zaznamenáno 19 806 mutací, ale mutace 124STOP mezi nimi není. Jedná se pravděpodobně o zcela novou mutaci, která dosud nebyla popsána.
Závěry
Potvrdili jsme, že FASAY je velmi spolehlivá metoda pro stanovení funkčního stavu proteinu p53. U buněčných linií, jejichž status p53 byl popsán již dříve, jsme předchozí výsledek buď potvrdili nebo zpřesnili. V několika případech nám FASAY poskytla hlubší vhled do povahy statutu p53.
- Jako první jsme určili status p53 u buněčných linií ML-2,
MONOMAC-1, HPB-ALL.
- Detekovali jsme novou, dosud nepublikovanou „non-sense“ mutaci
p53 v kodónu 124. Byla nalezena v buňkách linie HPB-ALL.
- U linií SK-LMS-1 a SK-UT-1 jsme zaznamenali přítomnost dvou
bodových mutací p53. Metodou FASAY jsme prokázali, že pokud ke
ztrátě heterozygotnosti (LOH) u genu p53 dojde mechanismem druhé
bodové mutace, může se v takto postižených nádorech vyskytovat malá
frakce buněk, které nesou funkční alelu p53. Ta vzniká
pravděpodobně rekombinací mezi oběma alelami postiženými bodovými
mutacemi. Frekvence je závislá na vzdálenosti mutací. Tento
výsledek má praktickou relevanci, protože nádory s dvěma
nezávislými bodovými mutacemi p53 byly již nalezeny (5).
- Nalezli jsme nesrovnalost ve statutu p53 buněčné linie ML-1. V
závislosti na zdroji této linie jsme určili status p53 buď jako
heterozygotní (a) (MZLU, Brno) nebo funkční (b) (Oddělení
patologie, SUNY Stony Brook, USA). Tento nález ukazuje na možnost
kontaminace nebo záměn buněčných linií nebo na případný vznik nové
mutace p53 během kultivace buněk v tkáňové kultuře.
- Analýza buněčných linií Hs913T, HL60 a Saos-2 ukázala na
některé limity metody FASAY, která nedokáže detekovat vět-
ší přestavby nebo delece genu p53.
Tabulka 1 Přehled analyzovaných buněčných linií a jejich status p53 určený metodou FASAY (ALL – akutní lymfatická leukémie; ca. – karcinom; CML – chronická myeloidní leukémie; AML – akutní myeloidní leukémie; PML – promyelocytární leukémie; LMS – leiomyosarkom; APL – akutní promyelocytární leukémie).
Tabulka 2 Přehled nově detekovaných mutací p53 u vybraných buněčných linií.
Literatura
- Flaman JM, Frebourg T, Moreau V, Charbonnier F, Martin C,
Chappuis P, Sappino AP, Limacher JM, Bron L, Benhattar J, Tada M,
Van Meir EG, Estreicher A, Iggo RD. Proc.Natl.Acad.Sci.USA 92
(1995) 3963-3967.
- Šmardová J, Šmarda J, Koptíková J. Biol.listy 69 (2004)
191-214.
- Resnick M, Inga A. Proc.Natl. Acad.Sci.USA 100 (2003)
9934-9939.
- Hainaut P, Soussi T, Shomer B, Hollstein M, Greenblatt M, Hovig
E, Harris CC, Montesano R. Nucl.Acid Res. 25 (1997)
151-157.
- Šmardová J, Kšicová K, Binková H, Krpenský A, Pavlová Š,
Rottenberg J, Koukalová H. Oncol.Rep. 11 (2004) 923-929.
Práce byla podporována grantem IGA MZ ČR č. NR/8068-3.
- Flaman JM, Frebourg T, Moreau V, Charbonnier F, Martin C,
Chappuis P, Sappino AP, Limacher JM, Bron L, Benhattar J, Tada M,
Van Meir EG, Estreicher A, Iggo RD. Proc.Natl.Acad.Sci.USA 92
(1995) 3963-3967.
Datum přednesení příspěvku: 26. 5. 2005